Грибы — древние гетеротрофные организмы, занимающие особое место в общей системе живой природы. Они могут быть как микроскопически малы, так и достигать нескольких метров. Поселяются на растениях, животных, человеке или на мёртвых органических остатках, на корнях деревьев и трав. Их роль в биоценозах велика и разнообразна. В цепи питания они являются редуцентами — организмами, питающимися мёртвыми органическими остатками, подвергающими эти остатки минерализации до простых органических соединений.
В природе грибы играют положительную роль: они пища и лекарства для животных; образуя грибокорень, помогают растениям всасывать воду; являясь компонентом лишайников, грибы создают среду обитания для водорослей.
Грибы — бесхлорофилльные низшие организмы, объединяющие около 100 000 видов, от мелких микроскопических организмов до таких великанов, как трутовики, гигантский дождевик и некоторые другие.
В системе органического мира грибы занимают особое положение, представляя отдельное царство, наряду с царствами животных и растений. Они лишены хлорофилла и поэтому требуют для питания готовое органическое вещество (принадлежат к гетеротрофным организмам). По наличию в обмене мочевины, в оболочке клеток — хитина, запасного продукта — гликогена, а не крахмала — они приближаются к животным. С другой стороны, способом питания (путём всасывания, а не заглатывания пищи), неограниченным ростом они напоминают растения.
Грибы имеют и признаки, свойственные только им: почти у всех грибов вегетативное тело представляет собой грибницу, или мицелий, состоящий из нитей — гиф.
Это тонкие, как нити, трубочки, заполненные цитоплазмой. Нити, составляющие гриб, могут туго или рыхло переплетаться, ветвиться, срастаться друг с другом, образуя плёнки наподобие войлока или видимые простым глазом жгуты.
У высших грибов гифы разделены на клетки.
В клетках грибов может быть от одного до нескольких ядер. Кроме ядер, в клетках имеются и другие структурные компоненты (митохондрии, лизосомы, эндоплазматическая сеть и пр.).

Строение
Тело подавляющего большинства грибов построено из тонких нитчатых образований — гиф. Совокупность их образует грибницу (или мицелий).

Разветвляясь, мицелий образует большую поверхность, что обеспечивает всасывание воды и питательных веществ. Условно грибы делятся на низшие и высшие. У низших грибов гифы не имеют поперечных перегородок и мицелий представляет собой одну сильно разветвлённую клетку. У высших грибов гифы разделены на клетки.
Клетки большинства грибов покрыты твёрдой оболочкой, её нет у зооспор и вегетативного тела некоторых простейших грибов. В цитоплазме гриба содержатся структурные белки и не связанные с органоидами клетки ферменты, аминокислоты, углеводы, липиды. Органоиды: митохондрии, лизосомы, вакуоли, содержащие запасные вещества — волютин, липиды, гликоген, жиры. Крахмала нет. В клетке гриба имеется одно или несколько ядер.
Размножение
У грибов различают вегетативное, бесполое и половое размножение.
Вегетативное
Размножение осуществляется частями мицелия, специальными образованиями — оидиями (образующимися в результате распадения гиф на отдельные короткие клетки, каждая из которых даёт начало новому организму), хламидоспорами (образуются примерно так же, но имеют более толстую тёмноокрашенную оболочку, хорошо переносят неблагоприятные условия), путём почкования мицелия или отдельных клеток.
Для бесполого вегетативного размножения специальные приспособления не нужны, но потомков появляется не много, а мало.
При бесполом вегетативном размножении клетки нити, ничем не отличаются от соседних, вырастают в целый организм. Иногда, животные или движение среды разрывают гифу на части.

Бывает при наступлении неблагоприятных условий нить сама распадается на отдельные клетки, каждая из которых может вырасти в целый гриб.
Порой на нити образуются наросты, которые разрастаются, отпадают и дают начало новому организму.

Часто некоторые клетки наращивают толстую оболочку. Они могут выдерживать высыхание и сохраняют жизнеспособность до десяти и более лет, а в благоприятных условиях прорастают.

При вегетативном размножении ДНК потомков не отличается от ДНК родителя. При таком размножении не нужны специальные устройства, но количество потомков невелико.
Бесполое
При бесполом споровом размножении нить гриба образует специальные клетки, создающие споры. Эти клетки выглядят как веточки, неспособные расти и отделяющие от себя споры, или как крупные пузыри, внутри которых образуются споры. Такие образования называют спорангиями.
При бесполом размножении ДНК потомков не отличается от ДНК родителя. На образование каждой споры тратится меньше веществ, чем на одного потомка при вегетативном размножении. Бесполым путём одна особь производит миллионы спор, поэтому у гриба больше шансов оставить потомство.
Половое
При половом размножении появляются новые сочетания признаков. При этом размножении ДНК потомков образуется из ДНК обоих родителей. У грибов объединение ДНК происходит по-разному.
Разные способы обеспечить объединение ДНК при половом размножении грибов:


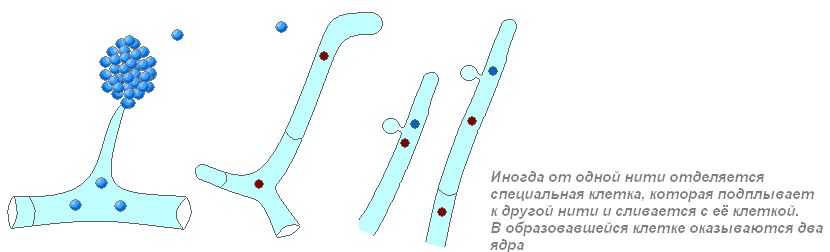
В какой-то момент сливаются ядра, а затем и нити ДНК родителей, обмениваются кусочками ДНК и разделяются. В ДНК потомка оказываются участки, полученные от обоих родителей. Поэтому потомок чем-то похож на одного родителя, а чем-то — на другого. Новое сочетание признаков может уменьшить, и увеличить жизнеспособность потомства.
Размножение состоит в слиянии мужских и женских половых гамет, в результате чего образуется зигота. У грибов различают изо-, гетеро- и оогамию. Половой продукт низших грибов (ооспора) прорастает в спорангий, в котором развиваются споры. У аскомицетов (сумчатых грибов) в результате полового процесса образуются сумки (аски) — одноклеточные структуры, содержащие обычно 8 аскоспор. Сумки образующиеся непосредственно из зиготы (у низших аскомицетов) или на развивающихся из зиготы аскогенных гифах. В сумке происходит слияние ядер зиготы, затем мейотическое деление диплоидного ядра и образование гаплоидных аскоспор. Сумка активно участвует в распространении аскоспор.
Для базидиальных грибов характерен половой процесс — соматогамия. Он состоит в слиянии двух клеток вегетативного мицелия. Половой продукт — базидия, на которой образуются 4 базидиоспоры. Базидиоспоры гаплоидны, они дают начало гаплоидному мицелию, который недолговечен. Путём слияния гаплоидного мицелия образуется дикариотический мицелий, на котором образуются базидии с базидиоспорами.
У несовершенных грибов, а в некоторых случаях и у других половой процесс заменяется гетерокариозом (разноядерностью) и парасексуальным процессом. Гетерокариоз состоит в переходе генетически неоднородных ядер из одного отрезка мицелия в другой путём образования анастомозов или слияния гиф. Слияние ядер при этом не происходит. Слияние ядер после, перехода их в другую клетку называется парасексуальным процессом.
Нити гриба прирастают поперечным делением (вдоль клетки нити не делятся). Цитоплазма соседних клеток гриба составляет единое целое — в перегородках между клетками есть отверстия.

Питание
Большинство грибов имеет вид длинных нитей, всасывающих питательные вещества всей поверхностью. Грибы всасывают нужные вещества из живых и мёртвых организмов, из почвенной влаги и воды природных водоёмов.
Грибы выделяют наружу вещества, разрывающие молекулы органических веществ на такие части, которые гриб может впитать.

Но в определённых условиях организму полезнее быть нитью (как гриб), а не комочком (циста) как бактерия. Проверим, так ли это.
Проследим за бактерией и растущей нитью гриба. Крепкий раствор сахара показан коричневым цветом, слабый — светло-коричневый, вода без сахара — белым.

Можно сделать вывод: нитевидный организм, разрастаясь, может оказаться в местах богатых пищей. Чем длиннее нить, тем больше запас веществ, который насытившиеся клетки могут расходовать на рост гриба. Все гифы ведут себя, как части одного целого, и участки гриба, оказавшись в богатых пищей местах, питают весь гриб.
Плесневые грибы
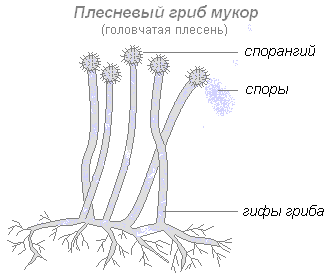
Плесневые грибы поселяются на увлажнённых остатках растений, реже животных. Одним из наиболее распространённых плесневых грибов является мукор, или головчатая плесень. Грибницу этого гриба в виде тончайших белых гифов можно обнаружить на залежавшемся хлебе. Гифы мукора не разделены перегородками. Каждая гифа представляет собой одну сильно разветвлённую клетку с несколькими ядрами. Одни ответвления клетки проникают в субстрат и поглощают питательные вещества, другие поднимаются вверх. На верхушке последних образуются чёрные округлые головки — спорангии, в которых образуются споры. Созревшие споры распространяются воздушными потоками или при помощи насекомых. Попав в благоприятные условия, спора прорастает в новую грибницу (мицелий).
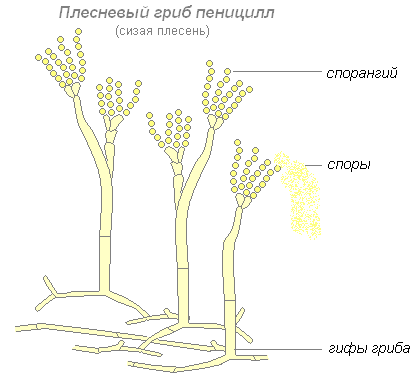
Вторым представителем плесневых грибов является пеницилл, или сизая плесень. Грибница пеницилла состоит из гифов, разделённых поперечными перегородками на клетки. Некоторые гифы поднимаются вверх, и на конце их образуются разветвления, напоминающие кисточки. На конце этих разветвлений образуются споры, с помощью которых пеницилл размножается.
Дрожжевые грибы
Дрожжи — одноклеточные неподвижные организмы овальной или удлинённой формы, размером 8-10 мкм. Настоящего мицелия не образуют. В клетке имеется ядро, митохондрии, в вакуолях накапливается много веществ (органических и неорганических), в них происходят окислительно-восстановительные процессы. Дрожжи накапливают в клетках волютин. Вегетативное размножение почкованием или делением. Спорообразование наступает после многократного размножения почкованием или делением. Оно совершается легче при резком переходе от обильного питания к незначительному, при поступлении кислорода. В клетке число спор парное (чаще 4-8). У дрожжей известен и половой процесс.
Дрожжевые грибы, или дрожжи, встречаются на поверхности плодов, на содержащих углеводы растительных остатках. От других грибов дрожжи отличаются тем, что не имеют грибницы и представляют одиночные, в большинстве случаев овальные клетки. В сахаристой среде дрожжи вызывают спиртовое брожение, в результате которого выделяются этиловый спирт и углекислый газ:
С 6 Н 12 О 6 → 2С 2 Н 5 ОН + 2СО 2 + энергия.
Этот процесс ферментативный, протекает при участии комплекса ферментов. Освобождающаяся энергия используется дрожжевыми клетками на жизненные процессы.
Размножаются дрожжи почкованием (некоторые виды — путём деления). При почковании на клетке образуется выпуклость, напоминающая почку.

Ядро материнской клетки делится, и одно из дочерних ядер переходит в выпуклость. Выпуклость быстро растёт, превращается в самостоятельную клетку и отделяется от материнской. При очень быстром почковании клетки не успевают разъединяться и в результате получаются короткие непрочные цепочки.
Не менее ¾ всех грибов — сапрофиты. Сапрофитный способ питания связан преимущественно с продуктами растительного происхождения (кислая реакция среды и состав органических веществ растительного происхождения более благоприятны для их жизни).
Грибы-симбионты связаны преимущественно с высшими растениями, мохообразными, водорослями, реже — с животными. Примером могут быть лишайники, микориза. Микориза — это сожительство гриба с корнями высшего растения. Гриб помогает растению усваивать труднодоступные вещества гумуса, способствует поглощению элементов минерального питания, помогает своими ферментами в углеводном обмене, активизирует ферменты высшего растения, связывает свободный азот. От высшего растения гриб, очевидно, получает безазотные соединения, кислород и корневые выделения, способствующие прорастанию спор. Микориза очень распространена среди высших растений, она не обнаружена лишь у осоковых, крестоцветных и водных растений.
Экологические группы грибов
Почвенные грибы
Почвенные грибы участвуют в минерализации органического вещества, образовании гумуса и т.п. В этой группе выделяют грибы, попадающие в почву только в определённые периоды жизни, и грибы ризосферы растений, живущие в зоне их корневой системы.
Специализированные почвенные грибы:
- копрофиллы - грибы, обитающие на почвах, богатых перегноем (навозные кучи, места скопления помёта животных);
- кератинофиллы - грибы, обитающие на волосах, рогах, копытах;
- ксилофиты - грибы, разлагающие древесину, среди них различают разрушителей живой и мёртвой древесина.
Домовые грибы
Домовые грибы — разрушители деревянных частей построек.
Водные грибы
К ним относится и группа микоризных грибов-симбионтов.
Грибы, развивающиеся на промышленных материалах (на металле, бумаге и изделиях из них)
Шляпочные грибы
Шляпочные грибы поселяются на богатой перегноем лесной почве и из неё получают воду, минеральные соли и некоторые органические вещества. Часть органических веществ (углеводы) они получают от деревьев.
Грибница — главная часть каждого гриба. На ней развиваются плодовые тела. Шляпка и ножка состоят из плотно прилегающих друг к другу нитей грибницы. В ножке все нити одинаковы, а в шляпке они образуют два слоя — верхний, покрытый кожицей, окрашенной разными пигментами, и нижний.

У одних грибов нижний слой состоит из многочисленных трубочек. Такие грибы называют трубчатыми. У других нижний слой шляпки состоит из радиально расположенных пластинок. Такие грибы называют пластинчатыми. На пластинках и на стенках трубочек образуются споры, с помощью которых грибы размножаются.

Гифы грибницы оплетают корни деревьев, проникают в них и распространяются между клетками. Между грибницей и корнями растений устанавливается полезное для обоих растений сожительство. Гриб снабжает растения водой и минеральными солями; заменяя на корнях корневые волоски, дерево уступает ему часть своих углеводов. Только при такой тесной связи грибницы с определёнными породами деревьев возможно образование плодовых тел у шляпочных грибов.
Образование спор
В трубочках или на пластинках шляпки образуются особые клетки — споры. Созревшие мелкие и лёгкие споры высыпаются, их подхватывает и разносит ветер. Разносят их насекомые и слизни, а также белки и зайцы, поедающие грибы. Споры не перевариваются в пищеварительных органах этих животных и выбрасываются наружу вместе с помётом.
Во влажной, богатой перегноем почве споры грибов прорастают, из них развиваются нити грибницы. Грибница, возникающая из одной споры, может образовывать новые плодовые тела лишь в редких случаях. У большинства видов грибов плодовые тела развиваются на грибницах, образованных слившимися клетками нитей, берущих начало от разных спор. Поэтому клетки такой грибницы двухъядерные. Грибница растёт медленно, лишь накопив запасы питательных веществ, она образует плодовые тела.
Большинство видов этих грибов — сапрофиты. Развиваются на перегнойной почве, отмерших растительных остатках, некоторые на навозе. Вегетативное тело состоит из гиф, образующих находящуюся под землёй грибницу. В процессе развития на грибнице вырастают зонтикоподобные плодовые тела. Пенёк и шляпка состоят из плотных пучков нитей грибницы.
У части грибов на нижней стороне шляпки от центра к периферии радиально расходятся пластинки, на которых развиваются базидии, а в них споры — это гименофор. Такие грибы называют пластинчатыми. У отдельных видов грибов имеется покрывало (плёночка из неплодных гиф), защищающее гименофор. При дозревании плодового тела покрывало разрывается и остаётся в виде бахромы по краям шляпки или кольца на ножке.
У некоторых грибов гименофор имеет трубчатую форму. Это трубчатые грибы. Их плодовые тела мясистые, быстро загнивают, легко повреждаются личинками насекомых, поедаются слизнями. Размножаются шляпочные грибы спорами и частями мицелия (грибницы).
Химический состав грибов
В свежих грибах вода составляет 84-94% общей массы.
Белки грибов усваиваются только на 54-85% — хуже, чем белки других растительных продуктов. Усвоению препятствует плохая растворимость белков. Жиры, углеводы усваиваются очень хорошо. Химический состав зависит от возраста гриба, его состояния, вида, условий произрастания и др.
Роль грибов в природе
Многие грибы срастаются с корнями деревьев и трав. Их сотрудничество взаимовыгодно. Растения дают грибам сахар и белки, а грибы разрушают находящиеся в почве мёртвые остатки растений и всасывают всей поверхностью гиф воду с растворёнными в ней минеральными веществами. Корни, сросшиеся с грибами, называют микоризой. Большинство деревьев и трав образуют микоризу.
Грибы играют в экосистемах роль разрушителей. Они уничтожают мёртвую древесину и листья, корни растений и трупы животных. Все мёртвые остатки они превращают в углекислый газ, воду и минеральные соли — в то, что могут усвоить растения. Питаясь, грибы набирают вес и становятся пищей животных и других грибов.
(настоящие грибы)
высшие грибы – отдел грибов
✎ Что такое высшие грибы?
Высшие грибы или настоящие грибы характеризуются многоклеточным мицелием (или грибницей), а ещё, наличием в оболочках целлюлозы (клетчатки - главного строительного материала в растительном мире) и хитина (природного соединения из группы азотосодержащих полисахаридов, из которого состоит верхний кутикулярный покров у жуков), отсутствием спорангиев (органов, производящих споры) при бесполом размножении, которое происходит у них только с помощью конидий (неподвижных спор бесполого размножения).
Высшие грибы , объединённые иногда в группу Carpomycetes , с входящими в неё классами базидиомицеты (лат. Basidiomycetes) и аскомицеты (лат. Ascomycetes), и бо́льшей половиной несовершенных грибов, составляющих группу Fungi imperfecti , с входящим в неё классом дейтеромицеты (лат. Deuteromycetes), образуют плодовые тела, в противоположность низшим грибам, у которых, плодовые тела отсутствуют.
✎ Правила классификации высших грибов
По размерам все высшие грибы можно разделить на:
макромицеты (и относящаяся к ним группа гастеромицеты) - это макро скопические грибы, те есть - большие грибы, с крупными плодовыми телами, основными представителями которых являются грибы из 2-х основных классов грибов: базидиомицеты и аскомицеты (именно их собирают, заготавливают впрок и употребляют в пищу);
микромицеты - это микро скопические грибы, типичными представителями которых могут являться только грибы класса эндомицеты (лат. Endomycetes) (их вообще не собирают и никогда не используют в пищу, но тем не менее, их с большим успехом применяют в медицине).
Именно из-за такого разделения грибов на макромиценты и микромицеты, сначала и были приняты правила систематизации грибов в отделы по их размерности, и поэтому до недавних пор класс эндомицеты был составной частью отдела низших грибов.
А сейчас (в наше время) все грибы систематизируются по отделам на основе способов их размножения, поэтому класс эндомицетов занял место в отделе высших грибов. Но, если рассматривать эндомицеты не как самостоятельный подотдел высших грибов, а всего лишь как выделенный подкласс из класса аскомицетов, то это тоже грубой ошибкой не будет.
✎ Подотдел высших базидиальных грибов
Этот подотдел высших грибов состоит из 3-х классов:
-
* базидиомицеты (лат. Basidiomycetes),
* урединиомицеты (лат. Urediniomycetes),
* устилягиномицеты (лат. Ustilaginomycetes),
наибольший практический интерес из которых представляет только первый.
Есть несколько версий классификации базидиомицетов, но наиболее полная и подробная версия их классификации выглядит следующим образом:
трубчатые (пористые) болетовые грибы
" " " род болет (лат. Bolet)
" " " род обабок (лат. Leccinum)
" " " род моховик (лат. Xerocomus)
" " " род порфировик (лат. Porphyrellus)
" " " род тилопил (лат. Tylopilus)
" " семейство маслёнковые (лат. Suillaceae)
" " " род маслёнок (лат. Suillus)
" " семейство гиропоровые (лат. Gyroporaceae)
" " " род гиропор (лат. Gyroporus)
" " семейство шишкогрибовые (лат. Strobilomycetaceae)
" " " род шишкогриб (лат. Strobilomyces)
пластинчатые (пластинковые) болетовые грибы
" " семейство болетовые (лат. Boletaceae)
" " " род филлопор (лат. Phylloporus)
" " семейство мокруховые (лат. Gomphidiaceae)
" " " род мокруха (лат. Gomphidius)
" " " род хроогомфус (лат. Chroogomphus)
" " семейство свинушковые (лат. Paxillaceae)
" " " род свинушка (лат. Paxillus)
" " семейство тапинелловые (лат. Tapinellaceae)
" " " род тапинелла (лат. Tapinella)
пластинчатые (пластинковые) грибы
" " семейство мухоморовые (лат. Amantaceae)
" " " род мухомор (лат. Amanita)
" " " " секция поплавок (лат. Amanitopsis)
" " " род кататрама (лат. Catatrama)
" " " род лимацелла (лат. Limacella)
" " " род торрендия (лат. Torrendia)
" " семейство плютеевые (лат. Pluteaceae)
" " " род вольвариелла (лат. Volvariella)
" " " род плютей (лат. Pluteus)
" " семейство шампиньоновые (лат. Agaricaceae)
" " " род гриб-зонтик (лат. Macrolepiota)
" " " род чешуйница (лат. Lepiota)
" " " род белошампиньон (лат. Leucoagaricus)
" " " род цистодерма (лат. Cystoderma)
" " " род чернопластинник (лат. Melanophyllum)
" " " род шампиньон (лат. Agaricus)
" " " род головач (лат. Calvatia)
" " семейство навозниковые (лат. Coprinaceae)
" " " род навозник (лат. Coprinus)
" " " род лакримария (лат. Lacrymaria)
" " " род хруплянка (лат. Psathyrella)
" " " род панеолина (лат. Panaeolina)
" " семейство строфариевые (лат. Strophariaceae)
" " " род панеолюс (лат. Panaeolus)
" " " род ложноопёнок (лат. Hypholoma)
" " " род псилоцибе (лат. Psilocybe)
" " " род строфария (лат. Stropharia)
" " " род чешуйчатка (огнёвка) (лат. Pholiota)
" " семейство паутинниковые (лат. Cortinariaceae)
" " " род колпак (лат. Rosites)
" " " род волоконница (лат. Inocybe)
" " " род гебелома (лат. Hebeloma)
" " " род паутинник (лат. Cortinarius)
" " " род гимнопил (лат. Gymnopilus)
" " " род крепидотус (лат. Crepidotus)
" " " род кюнеромицес (лат. Kuehneromyces)
" " " род галерина (лат. Galerina)
" " " род феолепиота (лат. Phaeolepiota)
" " семейство энтоломовые (лат. Entholomotaceae)
" " " род энтолома (розовопластинник) (лат. Entholoma)
" " " род клитопилюс (лат. Clitopilus)
" " семейство рядовковые (лат. Tricholomataceae)
" " " род астерофора (лат. Asterophora)
" " " род белосвинушка (лат. (Leucopaxillus)
" " " род гигрофоропсис (лат. Hygrophoropsis)
" " " род говорушка (лат. Clitocybe)
" " " род денежка (лат. (Collybia)
" " " род калоцибе (лат. Calocybe)
" " " род лаковица (лат. Laccaria)
" " " род леписта (лат. Lepista)
" " " род лиофиллюм (лат. Lyophyllum)
" " " род меланолейка (лат. Melanoleuca)
" " " род мицена (лат. Mycena)
" " " род негниючник (лат. Marasmius)
" " " род омфалина (лат. Omphalina)
" " " род рядовка (лат. Tricholoma)
" " " род трихоломопсис (лат. Tricholomopsis)
" " " род фламмулина (лат. Phlammulina)
" " семейство вёшенковые (лат. Pleurotaceae)
" " " род вёшенка (лат. Pleurotus)
" " " род гоенбуелия (лат. Hohenbuelia)
" " " род нематоктонус (лат. Nematoctonus)
" " семейство больбитиевые (лат. Bolbitiaceae)
" " " род больбитий (лат. Bolbitius)
" " " род коноцибе (лат. Conocybe)
" " " род галеропсис (лат. Galeropsis)
" " семейство гигрофоровые (лат. Hygrophoraceae)
" " " род гигрофор (лат. Hygrophorus)
" " " род гигроцибе (лат. Hygrocybe)
" " семейство сыроежковые (лат. Russulaceae)
" " " род сыроежка (лат. Russula)
" " " род млечник (груздь) (лат. Lactarus)
трутовиковые (трутовые) грибы
" порядок герициевые (лат. Hericiales)
" " семейство герициевые (лат. Hericiaceae)
" " " род гериций (лат. Hericium)
" " " род креолофус (лат. Creolophus)
" " семейство пилолистниковые (лат. Lentinellaceae)
" " " род лентинус (лат. Lentinus)
" " семейство аурискальпиевые (лат. Auriscalpeaceae)
" " " род аурискальпиум (лат. Auriscalpium)
" " " род лентинеллус (лат. Lentinellus)
" порядок телефоровые (лат. Telephorales)
" " семейство телефоровые (лат. Telephoraceae)
" " " род полиозеллус (лат. Polyozellus)
" порядок стереумовые (лат. Stereales)
" " семейство стереумовые (лат. Stereceae)
" " " род стереум (лат. Stereum)
" " семейство мерулиусовые (лат. Meruliaceae)
" порядок гименохетовые (лат. Hymenochaetales)
" " семейство гименохетовые (лат. Hymenochaetaceae)
" порядок ганодермовые (лат. Ganodermatales)
" " семейство ганодермовые (лат. Ganodermataceae)
" " " род ганодерма (лат. Ganoderma)
" порядок пориевые (лат. Poriales)
" " семейство кориоловые (лат. Coriolaceae)
" " семейство полипоровые (лат. Polyporaceae)
" " " род полипорус (лат. Polyporus)
" " " род пикнопорус (лат. Pycnoporus)
" " " род криптопорус (лат. Cryptoporus)
" " " род фомес (лат. Fomes)
" " " род траметес (лат. Trametes)
" " " род ленцитес (лат. Lenzites)
" " " род переннипория (лат. Perenniporia)
" " " род пория (лат. Poria)
" порядок фистулиновые (лат. Fistulinales)
" " семейство фистулиновые (лат. Fistulinaceae)
" " " род печёночница (лат. Fistulina)
индефинитные (неопределённые) грибы
" порядок лисичковые (лат. Cantharellales)
" " семейство ежовиковые (лат. Hydnaceae)
" " " род ежовик (лат. Hydnum)
" " семейство лисичковые (лат. Cantharellaceae)
" " " род лисичка (лат. Cantharellus)
" " семейство вороночниковые (лат. Craterellaceae)
" " " род вороночник (лат. Craterellus)
" " семейство клавариадельфовые (лат. Clavariadelphaceae)
" " " род рогатик (лат. Clavariadelphus)
" " семейство клавариевые (лат. Clavariaceae)
" " " род булавница (лат. Clavaria)
" " семейство спарассисовые (лат. Sparassidaceae)
" " " род спарассис (лат. Sparassis)
" порядок гомфовые (лат. Gomphales)
" " семейство гомфовые (лат. Gomphaceae)
" " " род гомфус (лат. Gomphus)
" " " род клавариадельфус (лат. Clavariadelphus)
" " " род рамария (лат. Ramaria)
" порядок аурикуляриевые (лат. Auriculariales)
" " семейство аурикуляриевые (лат. Auriculariaceae)
" " " род аурикулярия (лат. Auricularia)
дрожалковые (анаморфные) грибы
" порядок дрожалковые (лат. Tremellales)
" " семейство дрожалковые (лат. Tremellaceae)
" " " род дрожалка (лат. Tremella)
" " " род криптококк (лат. Cryptococcus)
" " семейство эксидиевые (лат. Exidiaceae)
" " " род эксидия (лат. Exidia)
" порядок дакримицесовые (лат. Dacrymycetales)
" " семейство дакримицесовые (лат. Dacrymycetaceae)
" " " род калоцера (лат. Calocera)
нутревиковые (нутриевые) грибы - гастеромицеты
" порядок весёлковые (лат. Phallales)
" " семейство весёлковые (лат. Phallaceae)
" " " род весёлка (лат. Phallus)
" " " род мутинус (лат. Mutinus)
" " " род решёточник (лат. Clathrus)
" порядок гнездовковые (лат. Nidulariales)
" " семейство гнездовковые (лат. Nidulariaceae)
" " " род гнездовка (лат. Nidularia)
" " " род бокальчик (лат. Cyathus)
" " " род круцибулюм (лат. Crucibulum)
" порядок тулостомовые (лат. Tulostomatales)
" " семейство тулостомовые (лат. Tulostomataceae)
" " " род тулостома (лат. Tulostoma)
" " " род баттаррея (лат. Battarrea)
" порядок дождевиковые (лат. Lycoperdales)
" " семейство дождевиковые (лат. Lycoperdaceae)
" " " род дождевик (лат. Lycoperdon)
" " семейство звездовиковые (лат. Geastraceae)
" " " род звездовик (лат. Geastrum)
" порядок ложнодождевиковые (лат. Sclerodermatales)
" " семейство ложнодождевиковые (лат. Sclerodermataceae)
" " " род ложнодождевик (лат. Scleroderma)
" " " род красноустка (лат. Calostoma)
" " " род мириостома (лат. Myriostoma)
" " " род хоракиелла (лат. Horakiella)
" " " род пизолитус (лат. Pisolithus)
" " " род тремеллогастер (лат. Tremellogaster)
" " " род хлорогастер (лат. Chlorogaster).
✎ Подотдел высших сумчатых грибов
Этот подотдел высших грибов включает в себя около 10-и классов, наиболее значимым из которых считается класс аскомицетов (лат. Ascomycetes) - один из крупнейших классов грибов, в котором до 2-х тысяч родов и более 30-и тысяч видов, что составляет около 30-и % от всех известных видов грибов.
Аскомицеты (от греч. аskos - сумка и mykes - гриб) получили такое название из-за наличия аски (или сумки) - органа плодоношения, содержащего 4-е или 8-мь гаплоидных половых аскоспор. Гифы имеют выраженные перегородки, а размножаются они половым (через образование аскоспор) и бесполым (через формирование конидий) путём.
К аскомицетам относят:
- блюдцевики,
- сморчки,
- строчки,
- трюфели,
- лопастники,
и многие другие грибы, в том числе и грибы-дрожжи (класс сахаромицетов (лат. Saccharomycetes)) - простых одноклеточных грибов, утративших способность образовывать мицелий (грибницу).
На основе наличия или отсутствия плодовых тел и их типа, класс аскомицеты (лат. Ascomycetes) делят на три подкласса:
Первый подкласс (низших аскомицетов) - гемиаскомицеты (или же голосумчатые) (лат. Hemiascomycetidae) - примитивные аскомицеты, с отсутствующими у них плодовыми телами, а их сумки образуются непосредственно на мицелии и они - прототуникатные (то есть, их сумки имеют тонкую, не дифференцированную оболочку, которая разрушается или растворяется, освобождая аскоспоры пассивно). К этому подклассу относят 4-е порядка.
Второй подкласс (высших аскомицетов) - эуаскомицеты (лат. Euascomucetidae) и их аски (или сумки) образуются в клейстотециях (закрытого типа плодовых телах), перитециях (полузакрытого типа плодовых телах) или же апотециях (открытого типа плодовых телах) и они - прототуникатные или уникатные. К этому подклассу относят около 20-и порядков.
Третий - (высших аскомицетов) - локулоаскомицеты (лат. Loculoascomucetidae) - сумки образуются в аскостромах и они - битуникатные (их сумки окруженны двойной оболочкой). Ну, а в настоящее время, этот порядок чаще называют эуроциальными (лат. Eurotiales) или же аспергиллальными (лат. Aspergillales). Он объединяет около 130-и родов, в которые входит около 270-и видов.
Ориентировочная классификация аскомицетов выглядит следующим образом:
аскомицеты (сумчатые грибы)
** подкласс гемиаскомицеты (лат. Hemiascomycetidae)
" порядок эндомицетовые (лат. Endoniycetales)
" " семейство диподасковые (лат. Dipodascaceae)
" " семейство эндомицетовые (лат. Endomycetaceae)
" " семейство сахаромицетовые (лат. Saccharomycetaceae)
" " семейство спермофторовые (лат. Spermophthoraceae)
" порядок протомицетовые (лат. Protomycetales)
" " " род протомицес (лат. Protomyces)
" " " род тафридиум (лат. Taphridium)
" " " род протомикопсис (лат. Protomycopsis)
" " " род миксия (лат. Mixia)
" порядок аскосферовые (лат. Ascosphaerales)
" " " род аскосфера (лат. Ascosphaera)
" " " род ульевая беттсия (лат. Bettsia alvei)
** подкласс эуаскомицеты (лат. Euascomucetidae)
" порядок эуроциевые (аспергиловые) (лат. Eurotiales)
" " " род эупенициллиум (лат. Eupenicillium)
" " " род таларомицес (лат. Talaromyces)
" " " род эмерицеллопсис (лат. Emericellopsis)
" " " род монаскус (лат. Monascus)
** подкласс лакулоаскомицеты (лат. Loculoascomucetidae)
" порядок плектасковые (лат. Plectascales)
дискомицеты (сумчатые грибы)
** подкласс пецицомицетиды (лат. Pezizomycatedae)
" порядок пецицевые (лат. Pezizales)
" " семейство лопастниковые (лат. Helveltaceae)
" " " род лопастник (лат. Helvella)
" " семейство пецициевые (лат. Pezizaceae)
" " " род пецица (лат. Peziza)
" " семейство пиронемовые (лат. Pyronemataceae)
" " " род алеврия (лат. Aleuria)
" " " род антракобия (лат. Anthracobia)
" " " род биссонектрия (лат. Byssonectria)
" " " род геопиксис (лат. Geopyxis)
" " " род гумария (лат. Humaria)
" " " род лампроспора (лат. Lamprospora)
" " " род меластица (лат. Melastiza)
" " " род октоспора (лат. Octospora)
" " " род пиронема (лат. Pyronema)
" " " род скутеллиния (лат. Scutellinia)
" " " род тарзетта (лат. Tarzetta)
" " " род хейлимения (лат. Cheilymenia)
" " семейство саркосцифовые (лат. Sarcoscyphaceae)
" " " род саркосцифа (лат. Sarcoscypha)
" " " род виннея (лат. Wynnea)
" " семейство сморчковые (лат. Morcheltaceae)
" " " род сморчок (лат. Morchella)
" " " род шапочка (лат. Verpa)
" " " род блюдцевик (лат. Disciotis)
" " семейство дисциновые (лат. Discinaceae)
" " " род строчок (лат. Gyromitra)
" " " род дисцина (лат. Discina)
" " семейство трюфелевые (лат. Tuberaceae)
" " " род трюфель (лат. Tuber)
" " " род парадокса (лат. Paradoxa)
" " " род хойромицес (лат. Choiromyces)
** подкласс леотиомицетиды (лат. Leotiomycatedae)
" порядок гелоциевые (лат. Helotiales)
" " семейство аскокортициевые (лат. Ascocorticiaceae)
" " " род аскокортициум (лат. Ascocorticium)
" " семейство дерматеевые (лат. Dermateaceae)
" " семейство гелоциевые (лат. Helotiaceae)
" " семейство геоглоссовые (лат. Geoglossaceae)
" " семейство гиалосцифовые (лат. Hyaloscyphaceae)
" " " род белонидиум (лат. Belonidium)
" " " род дазисцифус (лат. Dasyscyphus)
" " " род трихопецицелла (лат. Trichopezizella)
" " семейство склеротиниевые (лат. Sclerotiniaceae)
" " семейство фацидиевые (лат. Phacidiaceae)
пиреномицеты (сумчатые грибы)
** подкласс сордаромицетиды (лат. Sordariomycatedae)
" порядок ксиляриевые (лат. Xylareales)
" " семейство амфисфериевые (лат. Amphisphaeriaceae)
" " семейство гипонектриевые (лат. Hyponectriaceae)
" " семейство графостромовые (лат. Graphostromataceae)
" " семейство диатриповые (лат. Diatrypaceae)
" " семейство иодосфериевые (лат. Iodosphaeriaceae)
" " семейство клипеосфериевые (лат. Clypeosphaeriaceae)
" " семейство ксиляриевые (лат. Xylariaceae)
" " семейство миелоспермовые (лат. Myelospermataceae)
" порядок гипокрейные (лат. Hepocreales)
" " семейство гипокрейные (лат. Нуросгеасеае)
" " " род гипокрея (лат. Нуросгеа)
" " " род подострома (лат. Podostroma)
" " " род микоцитрус (лат. Mycocilrus).
✎ Подотдел высших голосумчатых грибов
Голосумчатые (иное название - первичносумчатые) (лат. Hemiascomycetidae) - это наиболее древняя и примитивная группа, вполне возможно, исходная для всех аскомицетов, а поэтому-то она и выделена в отдельный подотдел высших грибов, которую некоторые принимают за подкласс сумчатых грибов, содержащий:
Подотдел тафриномицеты (лат. Taphrinomycotina)
* класс тафриномицеты (лат. Taphrinomycetes)
" порядок тафриновые (лат. Taphrinales)
" " семейство тафриновые (лат. Taphrinaceae)
" " " род тафрина (лат. Taphrina).
Грибы являются одним из наиболее сложных для систематики объектов, особенно для создания естественной, филогенетической системы. Научные представления о грибах, об их происхождении и месте в системе живого мира бурно развивались и часто менялись в течение всего периода изучения этих организмов, это отражалось и на систематике.
Все грибы делят на группы: низшие (мицелий без перегородок) и высшие (мицелий с перегородками).
Низшие грибы.
В эту группу входит около тысячи видов с нечленистым, одноклеточным мицелием . Иногда гифы у низших грибов не образуются, а возникает плазмодий - разрастание цитоплазмы с многими ядрами. Диплоидное состояние только в зиготе, вся жизнь проходит в гаплофазе.
Оомицеты
Развитый мицелий, в клеточной оболочке отсутствует хитин, но есть целлюлоза.
Обитают во влажной или водной среде.
Возбудитель болезни картофеля, томата

Зигомицеты
Грибница хорошо развита, большей частью не разделена поперечными перегородками на отдельные клетки, лишь органы размножения часто отчленены перегородками.
Высшие грибы.
У высших грибов
мицелий разделен перегородками на отдельные клетки, содержащие одно, два или много ядер.
У некоторых грибов, например дрожжей, вегетативное тело представлено одиночными почкующимися клетками. Если клетки при почковании не расходятся, из них образуются цепочки - псевдомицелий. Некоторые примитивные грибы имеют одноклеточный таллом, иногда лишенный клеточной стенки.
|
Отдел |
Краткое описание |
Представители |
|
|
Аскомицеты |
Для большинства характерно образование асков - сумок с гаплоидными спорами, образующимися в процессе мейоза. Аскомицеты представляют собой одну из самых многочисленных групп грибов — более 32000 видов (~30 % всех известных науке видов грибов). Их отличает огромное разнообразие — от микроскопических почкующихся форм до обладающих очень крупными плодовыми телами грибов. |
Дрожжи - одноклеточные грибы, способные в вегетативному размножению путем почкования (пекарские, винные, кормовые) |
Дрожжи
|
|
Дейтеромицеты, или Несовершенные грибы |
Сводная группа (класс) несовершенных грибов, не имеющих полового спороношения. Включает более 25 тыс. видов, образующих септированный мицелий. Размножаются дейтеромицеты бесполым путем с помощью конидий. Для человека патогенны представители родов криптококков, торулопсис, питироспорум, кандида, трихоспорум, родоторула, малассезиа, микроспорой, трихофитон, эпидермофитон, гистоплазма, бластомицетов, кладоспориум и др. |
Пеницилл Мицелий пенициллов в общих чертах не отличается от мицелия аспергиллов. Он бесцветный, многоклетный, ветвящийся. Основное различие между этими двумя близкими родами заключается в строении конидиального аппарата. У пенициллов он более разнообразен и представляет собой в верхней части кисточку различной степени сложности (отсюда его синоним «кистевик»). На основе строения кисточки и некоторых других признаков (морфологических и культуральных) в пределах рода установлены секции, подсекции и серии. |
|
|
Аспергилл - используется для получения лимонной, щавелевой кислот |
|||
|
Триходерма - вызывает кожные заболевания человека |
|

Кроме того, предполагают, что наличие гриба в тканях подземных органов могло также способствовать повышению устойчивости высшего растения к засыханию.[ ...]
Грибы. Грибы1 - это одна из самых больших и процветающих групп организмов. Разнообразие грибов охватывает такие организмы, как одноклеточные дрожжи, плесневые грибы, возбудители болезней и, наконец, высшие грибы, имеющие зачастую крупные размеры и употребляемые в пищу человеком.[ ...]
Особенно часто они встречаются в почве ризосферы многих травянистых растений. Многие из них вызывают заболевания высших растений.[ ...]
Грибы обладают весьма большой склонностью к симбиотическим образованиям: мицелий ряда грибов совместно с корнями высших растений формирует микоризу. Это позволяет создавать сложные трофические отношения, в процессе которых грибы приобретают возможность разлагать недоступные высшим растениям органические вещества почвы, способствовать растениям в усвоении фосфатов, соединений азота и вырабатывать вещества - «активаторы роста». В ответ они получают в первую очередь углеводы. Симбиоз в виде микоризы наиболее взаимопроникающий. Установлено, что при удалении гриба растения-хозяева замедляют рост, некоторые дают семена, которые не прорастают, что отмечено, в частности, у орхидей.[ ...]
У грибов этого рода мицелий астериноидный, темно-бурый, развивающийся на поверхности живых листьев, реже стеблей и ветвей высших растений. Плодовые тела кожистые, шаровидные, черно-бурые. Сумки почти шаровидные или яйцевидные, реже булавовидные.[ ...]
К высшим грибам относят классы сумчатых грибов и бази-диальных. Их вегетативное тело представляет собой хорошо развитый нитчатый многоклеточный мицелий, они обладают морфологически четко дифференцированными органами размножения. У сумчатых грибов бесполое вегетативное и половое размножение. Названы они так за то, что при половом размножении споры образуются в небольших овальной или продолговатой формы органах, названных сумками. Сумки заключены в плодовые тела - аскокарпы. Вегетативные споры, или конидии, формируются на дифференцированных отростках мицелия- конидиеносцах (рис. 24). Базидиальные грибы также размножаются половым и бесполым, или вегетативным, путем. В результате полового процесса образуются специальные органы - базидии, каждая из которых несет на себе четыре споры.[ ...]
Почвенные грибы и высшие растения находятся в тесной взаимосвязи. Своеобразным и достаточно благоприятным местообитанием для многих почвенных микроскопических грибов является ризосфера, т. е. слой почвы в 2-3 мм, непосредственно примыкающий к корням. Растение пропитывает ризосферный: слой почвы своими корневыми выделениями, содержащими различные энергетические вещества, представляющие прекрасный питательный субстрат для развития грибов. Помимо этого, ризосферный слой почвы насыщается корневым опадом, который также хорошо ассимилируется микроскопическими почвенными грибами. Кроме этого, корни растений механическим путем изменяют и разрыхляют структуру почвы, улучшая ее аэрацию. Поэтому в ризосфере обильно развиваются все почвенные микроорганизмы, в том числе и микроскопические грибы.[ ...]
Головневые грибы распространены от Арктики до тропиков, почти повсюду, где произрастают высшие растения, не исключая пустынь и гор.[ ...]
Последние нередко обозначаются как + и -.[ ...]
Трюфелевые грибы - обязательные микори-зообразователи и поэтому произрастают по соседству с теми или иными высшими растениями. Например, черные трюфели (Tuber melanospo-rum, Т. aestivum) растут в лесах с плюсконосными деревьями - дубом, буком, грабом, орешником. Эти деревья и особый тип почвы являются благоприятным условием для произрастания трюфелей. Белые трюфели (Т. magnatum, Choi-romyces meandriformis) растут в лиственных лесах с березой, тополем, ильмом, липой, ивой, рябиной, боярышником. Изредка трюфели образуют микоризу с такими деревьями, как можжевельник, пихта и сосна.[ ...]
Многие виды грибов находятся в симбиозе с высшими растениями и способствуют снабжению растений питательными веществами.[ ...]
У некоторых грибов (особенно у опенка и домовых грибов) тяжи более мощные, их называют ризоморфами (они достигают нескольких метров длиной и нескольких миллиметров толщиной). У ризоморфов стенки наружных гиф темного цвета, а внутренние гифы обычно белые. Назначение ризоморфов то же, что и тонких тяжей, причем в некоторых случаях внутри ризоморф имеются особые проводящие трубки - широкие гифы, напоминающие сосуды высших растений.[ ...]
Пути заражения высших растений головневыми грибами различны. Основных путей четыре. Первый характеризуется тем, что головневые споры, удерживающиеся на семенах или сохраняющиеся в почве, прорастают на ра-стении-хозяине, в частности на наклюнувшемся семени, еще до того, как образуются всходы. Этот путь заражения свойствен возбудителям твердой (мокрой) головни пшеницы (Tilletia caries, рис. 212), головни ржи (Tilletia secalis) и головни ячменя (Tilletia hordei), стеблевой головни ржи (Urocystis occulta) и стеблевой головни пшеницы (Urocystis tritici), пыльной головни, кукурузы (Sorosporium reilianum, табл. 51) и т. д.[ ...]
Пероноспоровые грибы чрезвычайно широко распространены на земном шаре. Некоторые из них встречаются решительно повсюду, где произрастают высшие растения. К ним относятся, например, цистоповые грибы, обнаруживаемые и за полярным кругом, и в тропиках. При продвижении с юга на север количество видов заметно уменьшается. Благодаря особенностям своего развития и образу жизни наибольшее распространение они имеют в низменностях, где поселяются на равнинах в стациях, примыкающих к рекам, озерам и другим естественным и искусственным водоемам, отличающихся особым микроклиматом, главным образом высокой влажностью. Многие виды обитают в предгорных зонах, отдельные встречаются и в горах, преимущественно в сырых ущельях на теневыносливых растениях. В нижнем горном поясе, примерно до 1000 м над уровнем моря, способны обитать грибы довольно большого числа видов. В высокогорьях - в средней, верхней и субальпийской зонах - их заметно меньше. Определенные равнинные виды не поднимаются в высокогорья, а высокогорные виды не спускаются в равнины. В степях и пустынях также обитает незначительное число видов. Однако и в пустыне зарегистрированы отдельные виды пероноспоровых грибов.[ ...]
Мицелий головневых грибов разрастается преимущественно в живых клетках растения-хозяина. Гифы похожи на тонкие бесцветные нити диаметром 5-8 мкм. В зараженных клетках гифы образуют гаустории, которые обычно имеют гроздевидную форму и способствуют обмену веществ между грибом и высшим растением.[ ...]
Химическое действие высших грибов на организм человека известно с давних пор. В Европе зловещую репутацию снискала бледная поганка (Amanita phalloïdes), и с веками эта репутация лишь упрочилась. В Северной Америке произрастают не менее опасные виды поганок, такие, как Amanita virosa, A, verna, A. tenuifolia и A. bisporigera: Отравление поганками тем опаснее, что его симптомы появляются лишь на поздней стадии, когда повреждения печени и почек уже необратимы и смертельный исход неминуем. О яде поганок мы знаем в основном благодаря блестящим исследованиям Теодора Виланда и Отто Виланда . Для поганок характерны две группы ядовитых веществ: фаллотоксины, представителем которых является фаллоидин (24, рис. 3), и аматоксины, например а-амани-тин (29). Для человека смертельная доза аманитина равна 0,1 мг/кг: стало быть, одной бледной поганки весом 50 г, содержащей около 7 мг этого яда, достаточно, чтобы убить человека. Это вещество, будучи введено в желудочно-кишечный тракт мыши в достаточно высокой дозе (5 мг/кг) и притом одновременно с ядом, полностью предохраняет это животное от отравления. Подобное наблюдение очень любопытно, однако терапевтическое использование антаманида ограничено тем, что он является лишь превентивным противо-яди ем.[ ...]
То, что у большинства высших растений называют корнями, на самом деле микориза («грибокорень») - тесный мутуализм грибов и корневой ткани, при котором грибы помогают хозяевам получать минеральное питание, а сами берут у растения часть необходимого им органического углерода (см. недавний обзор Harley, Smith, 1983). Лишь представители очень немногих семейств, например, крестоцветные, не образуют такой ассоциации. У большинства мхов, папоротников, плаунов, голосеменных и покрытосеменных обнаружены ткани, более или менее тесно переплетенные с мицелием грибов. Все доминанты основных типов растительности на Земле - лесные деревья, травы и кустарники - имеют хорошо выраженную микоризу. Ископаемые остатки древнейших наземных растений позволяют предполагать их тесную связь с грибами. У этих форм еще нет корневых волосков, а в некоторых случаях - даже корней, поэтому заселение суши могло зависеть от присутствия грибов-мутуалистов.[ ...]
Этот порядок сумчатых грибов объединяет несколько сотен видов, большинство из которых развивается на растительном опаде, засохших ветвях и листьях древесных растений, кустарников и кустарничков, а также на травянистых и высших споровых растениях. Некоторые грибы поражают зеленые органы растений, а также луб коры и являются причиной их отмирания, что часто приводит к массовой гибели молодых растений в питомниках, культурах и в лесу.[ ...]
Геном хлоропластов ряда высших растений состоит из 120 генов. Хлоропластный геном очень сходен с бактериальным геномом как по организации, так и по функциям. В митохондриальном геноме человека, вероятно, отсутствуют интроны, но в ДНК хлоропластов некоторых высших растений, а также в ДНК митохондрий грибов интроны обнаружены. Считают, что хлоропластные геномы высших растений остаются без изменений примерно несколько миллионов лет. Возможно, что такая древность характерна и для митохондриальных геномов млекопитающих, включая человека.[ ...]
Заражение растений головневыми грибами долгое время остается внешне незаметным. Гриб обычно проникает в очень молодые органы высшего растения, в дальнейшем рост этих органов и разрастание в них мицелия происходят одновременно. Многие головневые грибы, развиваясь в растении-хозяине, после прорастания головневых спор пронизывают весь его стебель, а также нередко и другие органы. Однако многие головневые грибы, например Ustilago maydis, не распространяются далеко от места прорастания споры. Заражение, происходящее подобным образом, местное.[ ...]
В настоящее время известно более 500 видов грибов из рода филлостикта. Они вызывают заболевания растений - филлостиктозы. Чаще они известны на представителях розоцветных, бобовых, сложноцветных и других семейств высших растений.[ ...]
Помимо всего перечисленного, процесс разложения отмерших грибов также создает в почве большой запас усвояемой формы пищи для высших растений. Надо иметь в виду, что число поколений микроскопических грибов в почве в зоне корней растений за вегетационный период сменяется от семи до десяти и иногда более раз.[ ...]
Второй том издания «Жизнь растений» описывает образ жизни грибов и близкой к ним группы организмов - слизевиков (миксомицетов). В популярной форме освещены общие сведения о строении грибов, способах их размножения, о типах спороношения и плодовых тел, о способах питания. Многообразие грибов описано в систематическом плане от низших к высшим Том иллюстрирован цветными и тоновыми вклейками, оригинальными рисунками, картами и схемами.[ ...]
Непосредственные эволюционные связи рода Суэ риэ с другими низшими грибами не установлены. Возможно, что переходные формы в процессе исторического развития выпали из филогенетического ряда. Характерное для грибов этого рода спороношение состоит из слоя тесно сплоченных конидиеносцев, напоминающего гимениальный слой у высших грибов, и у других низших грибов не встречается.[ ...]
Типичным примером симбиоза может служить тесное сожительство между грибами и водорослями, приводящее к образованию более сложного и более приспособленного к природным условиям растительного организма - лишайника. Другим ярким примером симбиотического сожительства в почве является симбиоз грибов с высшими растениями, когда грибы образуют на корнях растений м и-к о р и з у. Явно выраженный симбиоз наблюдается между клубеньковыми бактериями и бобовыми растениями.[ ...]
Недавно было высказано предположепие, что переходу водорослевого предка высших растений в условия наземного существования значительно способствовал симбиоз с грибами. Как известно, симбиоз с грибами характерен для большинства высших растений, причем наиболее обычной его формой является симбиоз грибов с подземными органами (так называемая микориза).[ ...]
Все разводимые виды и сорта винограда подвержены этому заболеванию (конечно, в разной степени). Этим грибом также поражаются почти все плодовые деревья, помимо этого круг растений-хозяев гриба охватывает розы, дуб, бобы, картофель, ель, сосну и другие растения. Наиболее сильно гриб развивается в сырых местах, чаще всего на глинистых и мергелистых почвах, переувлажненных. На таких почвах высшие растения бывают ослаблены из-за недостатка железа и марганца. Хотя общее количество железа может быть велико (так же как и марганца), эти элементы, особенно железо, находятся в форме двухвалентных ионов и не только недоступны для растения, но и оказывают токсическое действие. По склонам холмов, на аллювиальных песках, известковых и гранитных почвах заболевание растений наблюдается сравнительно реже.[ ...]
При повышенной температуре (больше 20° С) капли могут быстро высыхать, поэтому у некоторых видов питиевых грибов в процессе эволюции возникла способность к прорастанию отделившихся от зеоспорангиеносцев зооспорангиев в таких условиях непосредственно ростковой трубкой, которая внедряется в растительные ткани. Таким образом, в семействе питиевых в зависимости от условий обитания можно наблюдать все вариапты бесполого размножения, от типичных зооспорангиев, характерных для водорослей и водных грибов, до конидий, характерных для наземных высших грибов.[ ...]
Сформированные сумки плотно прилегают друг к другу, образуя более или менее правильный слой, имеющий сходство с гимением высших сумчатых грибов. Однако в отличие от последних гимений тафриновых грибов не заключен в какие-либо специальные плодовые тела. Слой сумок обычно имеет желтый, красный, розовый или фиолетовый оттенок, придающий необычную окраску зараженному органу. Диплоидное ядро в сумке после окончания ее формирования трижды делится. В результате этого деления возникают 8 гаплоидных ядер, которые дают начало 8 аскоспорам. Споры большей частью округлые или яйцевидные и достигают 7- 10 мкм в диаметре. Эти аскоспоры способны к почкованию, в результате чего число их в сумках может увеличиваться в 2-4 раза.[ ...]
Последние данные науки существенно расширяют наши представления о границах биосферы. Обнаружено, что споры бактерий и мицелий некоторых грибов не теряют жизнеспособности в условиях высокого вакуума (10 мм рт. ст.). Бактерии обнаружены в водах атомных реакторов, часть из них выдерживает облучение порядка 2-3 млн рад. При температурах жидкого воздуха, гелия, водорода ряд бактерий остаются живыми. Даже отдельные высшие растения и насекомые переносят температуру, приближающуюся к абсолютному нулю (-273,16°С). Живые бактерии, способные к размножению, обнаружены в нефтяных водах на глубине 2800 м, на дне океанов (11 км), в рассолах с концентрацией 250 г/л, при давлениях, равных 1000 атмосфер, после 5 суток кипячения, 150-250 лет анабиоза (Ковда). Тем не менее обычно считают, что биосфера как область жизни охватывает верхнюю часть литосферы, всю гидросферу, тропосферу и нижние слои стратосферы до 25 км в высоту. Повседневно реальные границы распространения живого являются более узкими.[ ...]
Целлюлоза - основная пища для этих организмов, и фермент необходим для ее переваривания. Имеются данные об образование целлюлазы также в высших растениях , где ее роль сводится, по-видимому, к размягчению клеточных стенок перед их ростом Для высших растений и большинства высших животных (кроме жвачных) целлюлоза не является питательным веществом. Поскольку целлюлоза нерастворима, ее необходимо расщеплять вне клеточной мембраны, т. е. на поверхности клетки гриба или на некотором расстоянии от нее. В местах контакта гиф грибов с клеточными стенками целлюлозных материалов образуются отверстия при этом растворение клеточных стенок наблюдается даже на некотором расстоянии от проникающих гиф . При культивировании грибы выделяют целлюлолитические ферменты в культуральную среду. О механизме выделения почти ничего неизвестно, хотя можно предположить, что секретируют живые клетки, а не мертвые.[ ...]
Митохондриями, или хондриосомами, называются органоиды клетки эукариотов, представляющие собой мембранные внутриклеточные образования. Форма и размеры их различны - от овальных и грушевидных телец до нитевидных или ветвистых. По своему назначению митохондрии представляют собой центры сосредоточения ферментов энергетического обмена. М. Н. Мейсель обнаружил , что клетки дрожжей при брожении содержат меньшее число митохондрий, которые гипертрофированы (бродильный тип клеток). Аналогичная картина наблюдается и в аэробных условиях при избытке углеводов в среде, особенно сахарозы и глюкозы. Цитологически наблюдается бродильная перестройка митохондриального аппарата.[ ...]
У микробиологов Советского Союза представление об акти-номицетах формировалось на основании взглядов, развитых крупнейшим советским микробиологом Н. А. Красильниковым, который не считал актиномицеты бактериями и выделял их в самостоятельную таксономическую категорию - класс актиномицетов. Красильников все лучистые грибы разделял на высшие и низшие. Организмы с хорошо развитым мицелием, имеющие органы размножения, отнесены к высшим. Организмы, не образующие мицелия, с клетками палочковидной или кокковидной формы были отнесены им к низшим. Приведенные здесь характерные черты актиномицетов являются по Красильникову признаками высших форм. Низшие - очень близки к грамположи-тельным бактериям. В 8-м издании определителя Берги все без исключения актиномицеты безоговорочно отнесены к бактериям.[ ...]
Все это указывает на глубокие изменения в почве после девятилетнего ежегодного применения гербицидов. В результате резко меняются условия существования микроорганизмов, количество и качество энергетического материала, поступающего в почву. Если в контроле приток свежего органического вещества обеспечивается за счет высших травянистых растений, то в варианте с гербицидами главным образом за счет опавшей хвои, мхов и некоторых видов синезеленых водорослей. Это обусловливает относительное снижение численности сапрофитных грибов и бактерий, использующих легкодоступные органические вещества, с одной стороны, и возрастание числа микроорганизмов, разрушающих гуматы, - с другой. Таким образом, происходят качественные сдвиги: доминирующее положение в сообществе почвенных микроорганизмов занимает автохтонная группировка, разлагающая почвенный перегной.[ ...]
Любопытно, что Колли /9/ уже в 1907 г. высказал предположение, что уксусная кислота является предшественником при образовании определенных растительных фенолоь. В 1935 г. Фишер /10/ отметил, что кислоты шикимовая и хинная могут служить промежуточными продуктами при образовании галловой кислоты. Опыты над микроорганизмами, плесенями и грибами, бактериальными мутантами, проведенные многими авторами за последние годы, подтверждают оба эти механизма. Доказательство того, что аналогичные процессы протекают и в высших растениях, основывается на принципе диалектического единства метаболических механизмов в природе. Результаты применения радиоизотопной техники в исследовании высших растений подтверждают вывод о таком единстве. В случае механизма образования фенольных соединений через шикимовую кислоту было установлено также ее присутствие и наличие в растительных тканях ее производных. Этот механизм подтверждается и участием в этой метаболической системе двух ферментов - 5-дегидро-хиназы и 5-дегидрошикиморедуктазы.[ ...]
Водоросли прямо или косвенно участвуют в обогащении почвы азотом. Многие сине-зеленые водоросли являются фиксаторами атмосферного азота. В почвах СССР обнаружено 95 видов водорослей, для которых экспериментально доказана азотофиксация. В целинных почвах умеренной полосы накопление азота водорослями достигает 17-24 кг ¡га, а на поливных полях тропической зоны - до 90 кг/га. Методом меченых атомов доказано, что фиксированный водорослями азот может усваиваться другими водорослями, грибами п высшими растениями.[ ...]
Жизнь каждого растения, как и животного, имеет начало и конец. В любом лесу наряду с живыми деревьями, кустарниками, травами есть бурелом, валежник, отломившиеся и упавшие на землю сучья и ветви. Везде бывает более или менее мощный слой подстилки, состоящей главным образом из опавших листьев, хвои и т. п. В полях, на лугах, в садах после периода вегетации растений также накапливается много растительных остатков. Вся эта масса органического вещества разлагается в основном под воздействием грибов, превращаясь в более простые соединения, а затем в почву. Таким образом грибы выполняют важную функцию в общем круговороте веществ в природе. Основная роль при этом принадлежит высшим базидиальным и сумчатым грибам. Но некоторые несовершенные грибы также принимают в этом участие.[ ...]
Подавляющее большинство одноклеточных организмов -существа бесполые и размножаются путем деления клетки, что ведет к непрерывному образованию новых особей. Деление прокариотической клетки, из которой, в основном, состоят эти организмы, начинается с деления митозом наследственного вещества -ДНК, вокруг половинок которой впоследствии образуются две ядерные области дочерних клеток - новых организмов. Поскольку деление происходит митозом, то дочерние организмы по наследственным признакам полностью воспроизводят материнскую особь. Многие бесполые растения (водоросли, мхи, папоротники), грибы и некоторые одноклеточные животные образуют споры - клетки с плотным и оболочками, защищающими их к неблагоприятных условиях средь!. При олагоприятных условиях ооолочка споры раскрывается и клетка начинает дслиться митозом, давая начало новому организму. Бесполым размножением является также почкование, когда от родительской особи отделяется небольшой участок тела, из которого затем развивается новый организм. Бесполым является также вегетативное размножение у высших растений. Во всех случаях при бесполом размножении воспроизводятся в больших количествах генетически идентичные организмы, практически полностью копирующие родительский организм. Для одноклеточных организмов клеточное деление - это акт выживания, так как организмы, которые не размножаются, обречены на вымирание. Размножение и связанный с ним рост вносят в клетку свежие материалы и эффективно препятствуют старению, сообщая тем самым ей потенциальное бессмерше.[ ...]
Актуальность исследования: Полиненасыщенные жирные кислоты (ПНЖК) представляют собой уникальный класс органических веществ, играющих важную роль в биологических системах. Исследования последних трех десятилетий вскрыли широкий спектр их функций в живых организмах. ПНЖК подвергаются биотрансформации липоксигеназами или циклооксигеназами, что приводит к образованию многочисленных низкомолекулярных регуляторов процессов, протекающих в клетках, тканях и организме в целом. Одной из самых важных ПНЖК является арахидоновая кислота (АК), которая выступает в роли непосредственного предшественника серии простагландинов, лейкотрие-нов и тромбоксанов. Основными областями применения АК являются: фармакология (предшественник различных лекарственных и профилактических препаратов применяемых при заболеваниях сердечно-сосудистой системы, печени и др.); косметическая промышленность (средства по уходу за кожей); пищевая промышленность (обогащение различных продуктов питания, в том числе искусственных детских молочных смесей и др.); сельское хозяйство (высокоэффективный стимулятор роста и защитных реакций растений) и др. В настоящее время основным источником получения АК являются липидные экстракты из печени свиньи и других органов животных, что делает их крупномасштабное производство неэффективным (содержание АК оставляет не более 0,2% в пересчете на сухую массу). В течение последних двадцати лет значительные успехи были достигнуты в области биотехнологического получения АК с помощью низших грибов и морских водорослей, которые в ряде случаев позволили осуществить ее промышленное производство. Однако существующие на сегодняшний день биотехнологии получения АК далеки от совершенства, поскольку ее выход в лабораторных условиях в лучших случаях составляет 13 г/л (Япония), а в среднем у различных исследователей около 6-г 10 г/л (Россия, США, Польша и др.). В связи с этим актуальным является поиск и создание отечественных продуцентов АК и эффективных биотехнологий ее получения на их основе. Данная работа была выполнена по планам важнейших НИР Уфимского государственного нефтяного технического университета (УГНТУ) в соответствии с федеральной целевой программой “Государственная поддержка интеграции высшего образования и фундаментальной науки” на 1997-2001 гг. и 2002-2006 гг.
(одноклеточные грибы)
низшие грибы – отдел грибов
✎ Что такое низшие грибы?
Низшие грибы
или одноклеточные грибы
, вегетативное тело которых образовано мицелием клеточного строения, обладают менее совершенным половым способом размножения, чем высшие грибы. У них сильно разветвлённый, несептированный (с отсутствием перегородок), многоядерный мицелий.
Все грибы объединяются в тот или иной класс, семейство и род, по внешнему сходству или наоборот, по отличию биологии вида и по особенности клеточного строения, физиологическим и биологическим свойствам. Но немаловажную, а может быть и ключевую роль в классификации грибов имеет тип их полового спороношения (или полового развития). Исходя из этого принципа, все низшие грибы помещены в 3-и основные класса:
-
* оомицеты (лат. Oomycetes);
* хитридиомицеты (лат. Chytridiomymycetes);
* зигомицеты (лат. Zygomymycetes).
Но, в дополнение к ним, в систематике низших грибов есть и ещё 3-и класса:
-
* гломеромицеты (лат. Glomeromymycetes);
* гифохитриомицеты (лат. Hyphochytriomymycetes);
* лабиринтуломицеты (лат. Labyrinthulomymycetes).
И в каждом из этих классов грибов есть опасные возбудители болезней.
✎ 1-й класс низших грибов
Оомицеты (от лат. оomycota или аlveolata - широкое корыто), в отличие например от хитридиомицетов, имеют хорошо развитый мицелий и половой процесс у них - оогамный, когда на мицелии образуются женские половые клетки (оогонии) и мужские половые клетки (антеридии). При их оплодотворении цитоплазма антеридия сливается с плазмой оогония и происходит слияние их ядер. Образовавшаяся при этом ооспора сохраняется в растительных тканях до наступления весны, когда и прорастает в мицелий. Но у некоторых видов ооспора образуется без оплодотворения. К оомицетам относят:
-
" порядок сапролегниевые (лат. Saprolegniales) - это:
сапрофитные грибки, которые живут в воде на разлагающихся животных или растительных веществах, их мицелий одноклеточный, ветвистый, нитевидный и бесцветный;
-
" порядок пероноспоровые (лат. Peronosporales) - это:
Представители этого класса грибов вызывают много опасных заболеваний, таких как: корнеед сахарной свеклы, фитофтороз картофеля, склероспороз многих культурных злаков (кукурузы, риса и проса). Одним из опаснейших врагов винограда является завезённый из Америки в Европу опаснейший гриб-патоген - плазмопара витикола, который может поражать все зелёные органы растений, вызывая мильдью (или ложную мучнистую росу) винограда - самую опасную грибную болезнь европейских сортов винограда, нанося громадный ущерб виноградарству. К оомицетам относятся также возбудители ложной мучнистой росы огурцов и салата, пероноспороза сахарной и столовой свеклы, махорки и табака и белой ржавчины крестоцветных.
✎ 2-й класс низших грибов
Хитридиомицеты (от греч. сhytridion - капелька) - это наиболее простые по строению микроорганизмы, развитие которых обязательно связано с водой. У некоторых хитридиомицетов вегетативное тело представляет амебоид (цитоплазматическую массу), а у других - мицелий (грибницу), состоящий из тончайших гиф (грибных нитей). И когда цитоплазматическая масса образует зооспорангий (одноклеточный орган бесполого размножения) с зооспорами, которые, при помощи жгутиков, передвигаясь поражают растение. Это и есть принцип бесполого размножения. При половом же размножении, в результате слияния ядер клеток, образуются так называемые цисты (покоящиеся мелкие споры), которые, после завершения периода покоя, делятся и из проросшей зиготы (клетки, образовавшейся в результате слияния гамет разного пола), выходят зооспоры. Таким образом, у них бесполое размножение сменяется половым.
Хитридиомицеты способны поражать молодые ткани и органы, вызывая у них гипертрофию (разрастание) и возникновение наростов на клубнях или корнях зелёных насаждений. Наиболее опасны среди них синхитриум эндобиотикум (лат. Synchytrium endobioticum) - возбудитель рака у картофеля и олпидиум брассика (лат. Olpidium brassicae) - возбудитель чёрной ножки у капусты.
✎ 3-й класс низших грибов
✎ Выводы и заключения
И тем не менее, не смотря на все негативные последствия, которые вызывает их жизнедеятельность, низшие грибы играют значимую роль в природе при разложении разного рода органических остатков и в подавлении численности многих вредных видов живых организмов.